Spatiotemporal
Patterns in Nanoclimate
And
Behavior
of the Lizard Gambelia wislizenii
Aliina Smith and
Advisor: Dr.
Abstract
A major goal in ecology is to document and
explain spatiotemporal patterns of distribution and abundance of organisms. The
first phase of ecological research is to observe and describe where individuals
are and when, with respect to environmental factors which motivate the organism. Desert scrub habitats are relatively simple, tractable
ecosystems and lizards are particularly tractable animals for ecological
studies in desert scrubs. Summer in the
This study was designed to examine the
spatiotemporal trends in 1) microclimate in Great Basin Desert Scrub, 2)
nanoclimate among microhabitats and nanohabitats of three predominant
mesohabitats of the
Insolation,
wind speed and air temperature within each mesohabitat in the study plot
followed a clear and predictable spatiotemporal pattern. Daily patterns of
direct solar radiation, substratum temperature in the open, and air temperature
all showed very similar temperature parabolas, differing only in the timing of
their peaks and troughs. Substratum temperature in the dappled/mixed shade
varied with time of day, microhabitat, and nanohabitat because of the continuous
change in the azimuth and declination of direct solar radiation, particularly
where the sunlight strikes beneath the foliage-perimeter of the shrubs. The data provided
by the copper models in this study can be directly applied towards predicting
and explaining G. wislizenii spatiotemporal behavior patterns. We can infer from the copper model
temperatures and other temperature measures that at both cool ends of the daily
activity period and during the heat of
Large
female G. wislizenii were relatively
and consistently less active throughout the day as compared with the other
size-sex classes. One explanation for
this activity difference may be differences among size-sex classes in modes of
food acquisition. Large female G.
wislizenii may use a long-wait ambush predation mode, spending relatively
long periods in the dappled lighting under shrub foliage, lying-in-wait to
ambush large, mobile prey such as lizards that may approach inadvertently. In contrast, smaller G. wislizenii may use more short-wait ambush predation mode wherein
they perform short visual scans of the foliage perimeters of shrubs, in search
of grasshoppers, cicadas, and other large arthropods that reveal themselves by
movement.
INTRODUCTION
There
are four basic ecological tasks or challenges that an animal faces: 1)
acquiring and utilizing food, 2) finding and acquiring mates, 3) avoiding and
evading predators, and 4) coping with abiotic stresses and avoiding abiotic
extremes. In deserts, animals face a
strong abiotic challenge via the spatiotemporal patterns of heat flow and the
effects of heat flow on body temperatures.
Thus, how the animals cope with heat flow, such as using physiological
and behavioral thermoregulation is especially important in small ectotherms
that live in deserts. Diurnally active
lizards, for example, behaviorally regulate their body temperatures within
narrow range of only a few degrees centigrade (Grant and Dunham, 1988; Muth,
1977). Thermoregulatory behaviors
include adopting specific postures that minimize or maximize exposure to solar
radiation and selecting nanohabitats at some periods of the day that permit body
temperatures to remain within a narrow range (Abts, 1976; Heath, 1965).
Primary
and venerable goals in ecological research are to document and explain
spatiotemporal patterns of individuals and populations. Observational-descriptive research of
individuals within and among populations is a necessary precursor to any
reliable experimental research. Desert lizards are particularly tractable field
research animals because in the sparse scrub vegetation, they are easy to find,
observe, and capture. The Long Nosed
Leopard Lizard (Gambelia wislizenii)
is an abundant, tractable lizard in the Great Basin Desert Scrub in the
HYPOTHESES
Given
that variation exists in all habitats, one would expect variation among the
mesohabitats of Great Basin Desert Scrub in their thermal spatiotemporal
patterns. Moreover, given that diurnally-active desert lizards thermoregulate
relatively precisely, one would expect variation in the thermoregulation
behavior of the desert-dwelling lizard Gambelia wislizenii to correlate
with this spatiotemporal variation in temperatures. Thus, this study examined
1) the spatiotemporal microclimate trends within the three predominant
mesohabitats of the
STUDY
SITE
The
study area is located in a 200 by 200 meter plot in the
MATERIALS
AND METHODS
Weather
station temperature measurements
For
one week in early July 2005, 24 thermocouples, attached to a multiplexer of a
portable weather station, were used to measure the temperatures of a variety of
nanohabitats in the sandy flat mesohabitat; minute-by-minute temperatures were
averaged once each 20 minutes (e.g., 0910 to 0930, 0930 to 0950, and 0950 to
1010 hrs). Prevalent, representative
microhabitats were used: two perennial plants Sarcobatus vermiculatus
and Artemisia tridentata plants, and open sandy flat areas. Thermocouple
probes were placed at the soil surface (substratum) in each cardinal direction,
and midway between the stem base of each perennial plant and the foliage
perimeter. Other thermocouples were placed: 25 cm above ground in the center
foliage of each plant, and both at the soil surface and 10 cm below ground just
north of the stem base in the presumptive coolest nanohabitats of the plant
microhabitat. Any thermocouple probe
that had a chance of being exposed to air was painted white to improve the albedo
and reduce heat absorption of direct sunlight by the probe.
Temperature
measurements with hand-held units
To
compare temperature trends among nanohabitats within a plant microhabitat and
to compare similar microhabitats between the dune and hardpan environments, a
medium-sized Greasewood (Sarcobatus vermiculatus) was selected for
temperature observations in each mesohabitat. Thermocouple probes were placed
in each cardinal direction at the base of each plant. Thermocouples wires were
prepared with the same method as the weather station thermocouple wires. Wires
were strung out to switch boxes situated in plastic containers and sheltered
with separate containers when not in use. Wind speed, air temperature, and
substratum temperatures were collected for each microhabitat in morning,
afternoon and evening time periods. The air temperature was determined by
attaching a single wire to the BARNANT 115 and maintaining the shaded
temperature probe at two meters above ground.
The substratum temperature was measured by placing the single wire
directly on the ground and covering it with a small patina of dust. Air and
substratum temperatures collected by researchers (with infrared, thermocouple,
and glass bulb mercury thermometers) on plot during other research activities
were used to buttress observations in this study. The substratum temperatures
measured with a cloacal thermometer were taken by gently resting the tip of the
thermometer against the substrate and dusting the tip with a few particles of
the local substratum. This process was repeated for measurements taken with the
Barnant 115 thermocouple thermometer. The non-contact infrared thermometer
measurements were taken by collecting readings from at least ten different
locations within a 0.25 meter radius on level ground. The highest and lowest
readings were removed (assuming inadvertent measures of substratum on tiny
slopes) and the remaining temperatures (assumed to be on level ground) were
then averaged and recorded.
iButton temperature measurements
Thermochron
iButtons were placed on the substratum in three microhabitat types across the
three predominant mesohabitats. The nanoclimates of the nanohabitats (weather
conditions of microhabitat or nanohabitats, as opposed to the weather
conditions of mesohabitats, typically known as the microclimates of
mesohabitats) of four large A. tridentata and four large S. vermiculatus
were selected for analysis with the iButtons. The specific iButton locations
(nanohabitats) in the plant microhabitats were 1) deep shade center of each
shrub, and 2) midway between the base and perimeter in each of the cardinal
directions of each shrub, and
the iButton in the open microhabitat, was placed on substratum surface
about 1 m south of each shrub. The iButtons were programmed to record
temperature once every ten minutes.
Copper
model temperature measurements
Six
copper models were constructed to reflect the three thermoregulation-relevant
body positions for both sexes of Gambelia
wislizenii, hence 3 models of males and 3 models of females. Copper models were based on average
snout-to-vent lengths (SVL) for each sex from a sample of the SVL measures of
least 30 adults of each sex taken the previous year. The female models were
constructed from copper tubing and glued together using epoxy. The male models
were constructed from aluminum tubing and were glued together with epoxy. Differences in heating between the copper and
aluminum tubing were considered negligible due to the fact that they both heat
up to the same final temperature. The males were constructed smaller than the
females in accordance with the previous year’s data. Models were glued together
using epoxy, spray-painted gray, and then hand-painted with a pattern of brown
spots to imitate the natural reflectance of G.
wislizenii. The thermocouple wire was inserted through a small hole drilled
where the vent would be found on an average lizard. Thermocouples wires were
prepared with the same method as the weather station thermocouple wires. Models
were used to test sunlit thermal variance between body positions (BROS = body
resting on substratum, FLEPOS = front legs extended, pelvis on substratum, and
ALEBNTS = all legs extended, body not touching substratum) and to test average
dappled/mixed shade temperature measurements of Sarcobatus vermiculatus microhabitat.
Focal
observations of Gambelia wislizenii behavior
G. wislizenii focal observations were
performed on 11 non-consecutive days in early July 2005, and observations were
confined to one of three time intervals each day. Morning observations were
made from
FIGURES
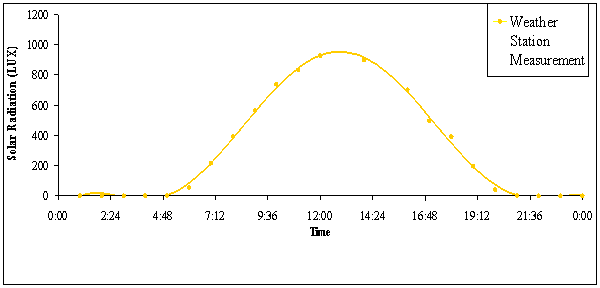
FIGURE 1. Hourly averages for intensity of solar
radiation on 5 days with clear skies in early July 2005
FIGURE 2. Average hourly wind speeds for 5 days in July
2005
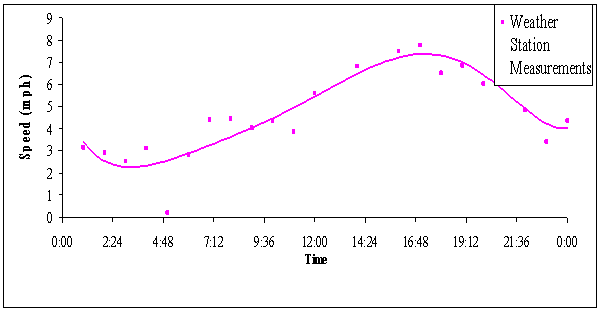
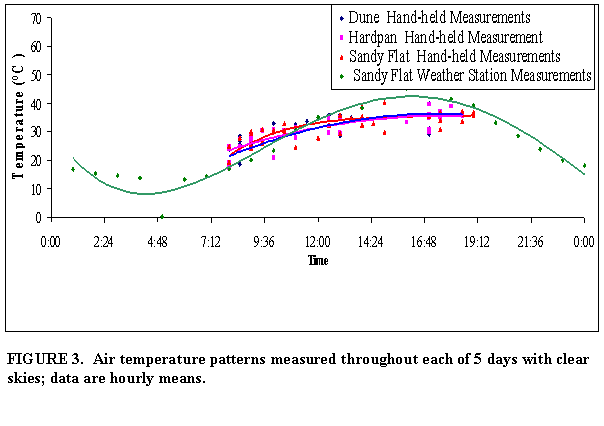
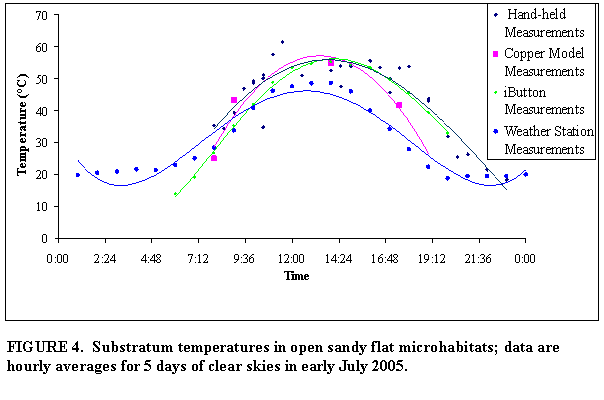
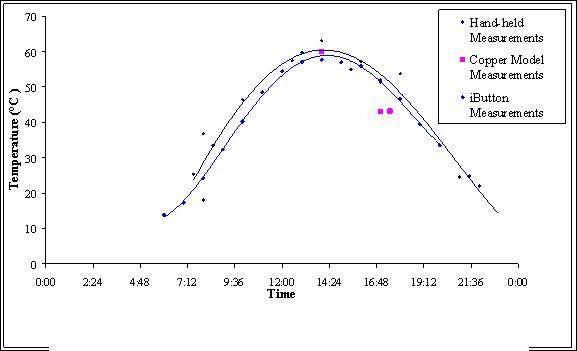
FIGURE 5. Substratum
temperatures in open dune microhabitats; data are hourly averages for 5
days of clear skies in early July 2005.
FIGURE 6. Substratum
temperatures in open harpan microhabitats; data are hourly averages for 5
days of clear skies in early July 2005.

Substratum north of S. vermiculatus root crown located
in the dune Substratum south of S. vermiculatus root crown located
in the dune
![]()
![]()
![]()
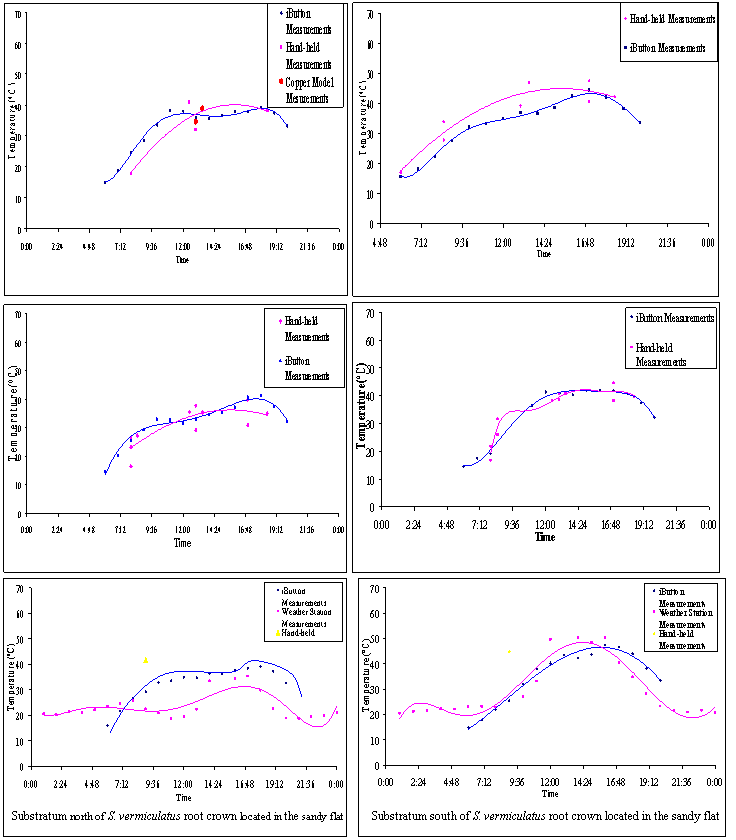
FIGURE 7. Hourly averages for substratum temperatures under a
mix of dappled shade and sun-shade edge of foliage of Sarcobatus
vermiculatus (in each mesohabitat) for 5 days of clear skies in early
July 2005.
![]()
FIGURE 8. Hourly averages for substratum temperatures under a
mix of dappled shade and sun-shade edge of foliage of Sarcobatus
vermiculatus (in each mesohabitat) for 5 days of clear skies in early July 2005.
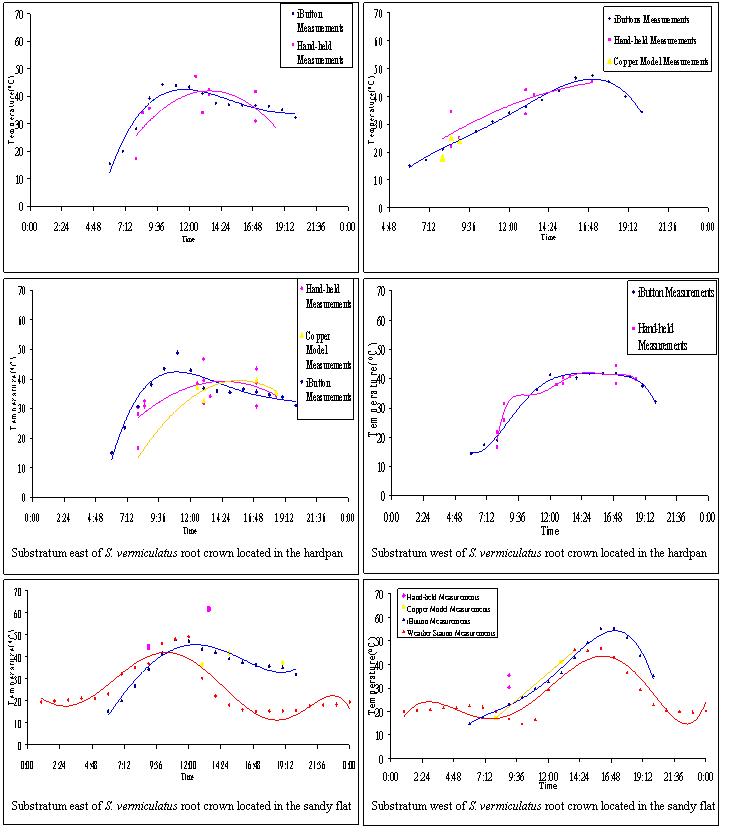
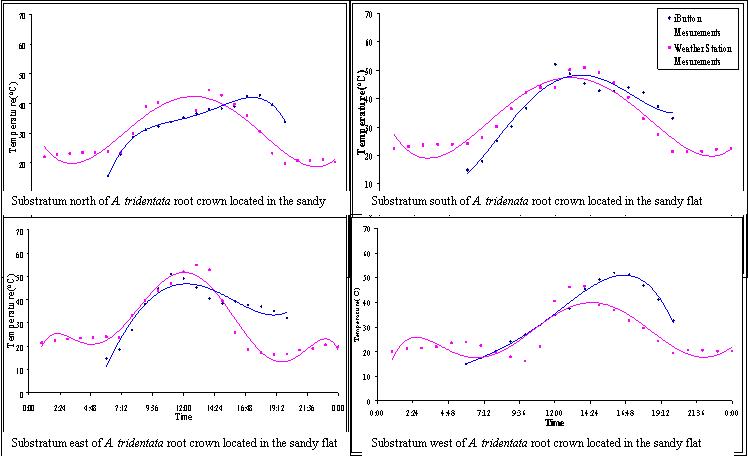
FIGURE 9.
Hourly averages
for substratum temperatures under a mix of dappled shade and sun-shade edge
of foliage of Artemisia tridentata (in sandy flats) for 5 days of
clear skies in early July 2005.
FIGURE 10. Hourly averages for temperatures of three nanohabitats
at the microhabitat of the perennial shrub Artemisia tridentata for
5 days with clear skies in early July 2005

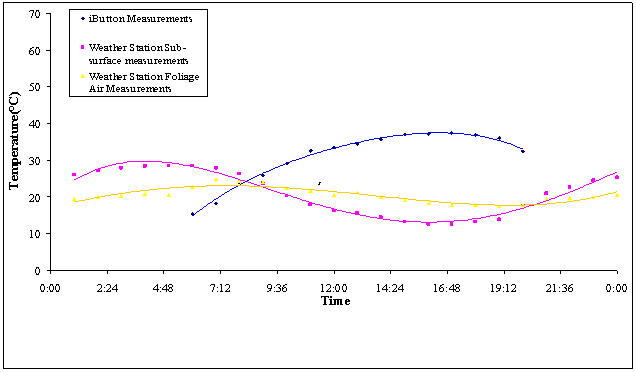
FIGURE 11. Hourly averages for temperatures of three nanohabitats
at the microhabitat of the perennial shrub Sarcobatus vermiculatus for
5 days with clear skies in early July 2005

FIGURE 12. Variation in
temperatures among copper lizard models of different a) lizard body positions,
b) mesohabitats, and c) microhabitats.
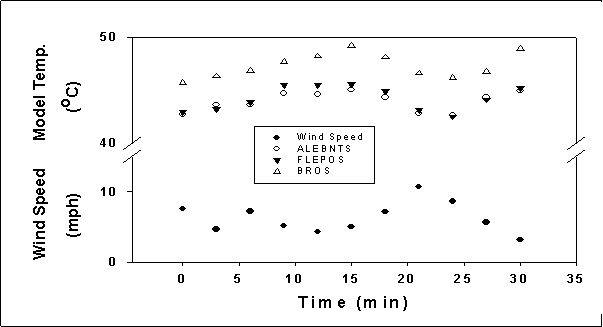
FIGURE 13. Comparisons of temperatures of copper lizard models of
different body positions as wind speed varies.
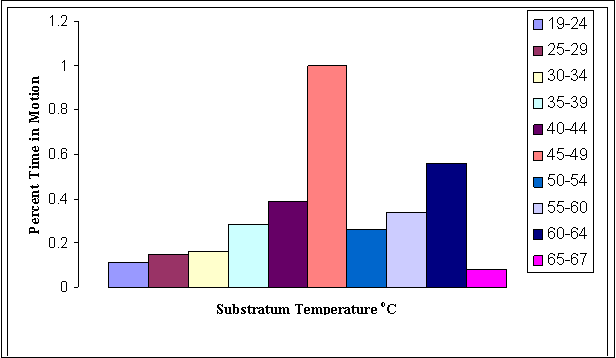
FIGURE 14. Proportion of
time spent moving by Gambelia wislizenii as related to sunlit
substratum temperatures in open microhabitat.
FIGURE 15. The inverse relationship of substratum
temperature in sunlit microhabitat with the proportion the total time
during focal observations that Gambelia
wislizenii were observed
to spend in that sunlit microhabitat.
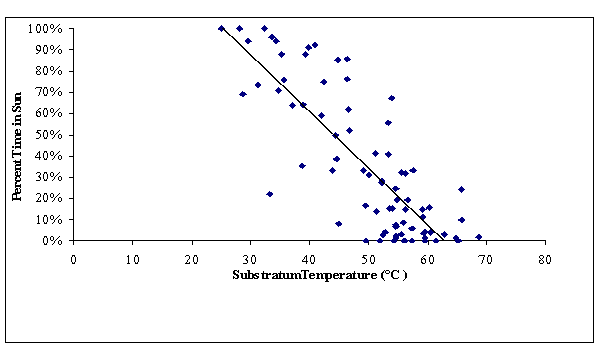

FIGURE 16. Body temperatures of
Gambelia wislizenii with respect to
time and mesohabitat (Hardpan temperatures appear to be
cooler in general, whereas sandy flat temperatures tend to be highest; note
that scarcity of lizards

FIGURE 17. The relative amount (%) of focal
observation time that large female Gambelia
wislizenii spent in motion as a function of sunlit substratum
temperatures in open microhabitats during mornings in early July 2005. (Note that the time spent moving appears to be
relatively stable and low.)
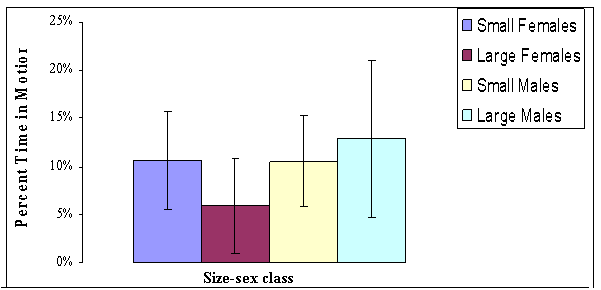
FIGURE 18. Comparisons among size-and-sex classes
of Gambelia wislizenii for how much time they spend moving
during morning (Data are shown
as average percent of observation time spent in motion for each size-and-sex
class from

FIGURE 19. Comparisons among size-and-sex classes of Gambelia wislizenii for how much time they spend moving
during
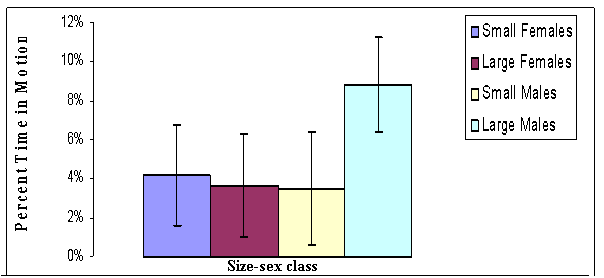
FIGURE 20. Comparisons among size-and-sex classes of Gambelia wislizenii for how much time they spend moving
during early evening (Data are
shown as average percent of observation time spent in motion for each size
class from
RESULTS
I.
Spatiotemporal microclimate trends within the three predominant
mesohabitats of the
Diurnal air
temperature, solar radiation and wind flow variation
Daily averages for
insolation for sequences of 20 min periods throughout the day ranged from 0 LUX
to 925 LUX, as measured by a pyranometer of an automated field weather station.
Solar radiation peaked approximately at
Average air
temperature for the sandy flat mesohabitat ranged from an average daily minimum
of 13.2oC before sunrise (
Wind speed recorded
by the weather station confirmed field personnel’s direct observations of low
wind in the morning and increased wind activity in the late afternoon. The
weather station recorded an average modal low of 2.5 mph at
Sunlit
substratum temperatures in open areas of sandy flat, hardpan and dune
Average temperature
trends for sunlit substratum in the sandy flat followed a characteristic
parabola, peaking near
Microclimate
of dappled/mixed shade at Sarcobatus vermiculatus
Putative soil
surface (substratum) temperatures measured by iButtons (essentially the
soil-air interface) within the north and south nanohabitats under the S. vermiculatus foliage (in the sandy
flats), with the iButtions placed midway between base and plant perimeter,
measured maximum temperatures about 1 hour later (6:00 hrs) than did either the
soil-surface thermocouple of the weather station or the various hand-held
temperature recording devices that measured substratum temperatures adjacent to
the copper models locations (Figure 8). Moreover, the single thermocouple probe
that measured substratum temperature for the weather station recorded lower
substratum temperatures than did the iButtons.
Another notable difference between iButton measurements and other
devices (weather station thermocouples and various hand-held units) was the
shape of the temperature curve. The
parabolic pattern of the iButton temperatures dropped slightly during the
Putative soil
surface (substratum) temperatures measured by iButtons within the east and west
nanohabitats under the S. vermiculatus
on all three mesohabitats displayed relatively little variation (44oC-48oC)
within mesohabitat type (Figure 9). The
west-side substratum under S.
vermiculatus located in the sandy flat, however, displayed a relatively
higher temperature peak of 58oC. In general, the temperatures were
slightly lower than the north-side and south-side substratum temperatures
relative to each mesohabitat type. The
nanohabitats of east-side and west-side substrata under the shade of foliage of
S. vermiculatus had noticeably
different temperature trends with respect to time. In contrast to north-side
and south-side iButtons, west-side iButtons recorded peak temperatures at
approximately the same time as the weather station/hand-held devices (
Comparisons of temperatures in two
microhabitats: Sarcobatus vermiculatus and Artemisia tridentata
In the sandy flat
mesohabitat, the putative soil surface (substratum) temperatures measured under
S. vermiculatus foliage midway
between base and plant perimeter and under A.
tridentata foliage midway between base and plant perimeter, displayed
relatively similar temperature trends, although A. tridentata nanohabitats were generally several degrees
centigrade warmer than the corresponding nanohabitats under S. vermiculatus (Figures 8 and 9). The
lag time between the other temperature gauges and iButton peak temperature
recordings in A. tridentata reflected
a similar timing with the north and south iButtons of S. vermiculatus. The
temperature patterns were virtually opposite of the S. vermiculatus east and west temperature parabolas, with a 1 hour lag time for westerly iButtons
and very little lag time for easterly (Figures 8 and 9).
Sub-surface soil
temperatures (-10cm) just north of the root-crown of S. vermiculatus in the
sandy flat peaked (28.6oC) at about 05:00hrs, and reached a trough
(12.5oC) at
II. Copper model operative environmental
temperatures
Averaged copper model
temperature measurements
Copper lizard model
temperatures placed in the open microhabitats of the sandy flat mesohabitat
reached 35oC by
Sunlit thermal variance
between copper model body positions
When comparing the
variances in temperature between the different body position models it is
apparent that in the open sunlit microhabitats, the models representing the
body position where the lizard has its body resting on the substratum (BROS)
become much warmer than either the FLEPOS (front legs extended and pelvis on
substratum) or the ALEBNTS model (all legs extended with body not touching the
substratum) (Figure 12). The open sunlit substratum reached the highest
temperatures among all microhabitats and nanohabitats, and all three models
showed distinctly different increasing temperature trends as substratum
temperatures rose. Differences among
models in temperature can be attributed to the amount of body contact with the
substratum. Note that the greater
average temperatures of the sandy flat areas produced the larger temperature
differences between model types (Figure 12).
The effects of the
wind on the models can also be seen in Figure 13. The BROS model, lying flat on
the ground was affected much less by the wind fluctuation in the sandy flat
areas, while the other two types of models showed similar variation between the
temperatures on sandy flat-open and sandy flat-protected (just leeward of
shrub) locations (Figure 13).
III. Focal observations of Gambelia wislizenii behavior
Although relatively few Gambelia wislizenii were seen at
The G. wislizenii decreased the amount of
time they spent in the sun as the substratum temperatures increased (Figure
15). This inverse relationship is
consistent with the findings of Tanner and Krogh (1974). Leopard lizard body
temperatures ranged lower in the hardpan areas than for dune and sandy flat
(Figure 16). Throughout the morning and evening time periods, lizard body
temperatures range from about 36.0 oC to as high as 42.0 oC.
Large female G. wislizenii seem to move at relatively
low and constant rates throughout the day (Figure 17). Their activity level is
generally lowest among size-sex classes (Figure 18, 19 and 20). Small females
apparently are more active than large females, and with respect to activity
levels, small females tend to behave more like small males. Small females,
however, are the most active size-sex class during the heat of
DISCUSSION
I.
Comparisons of spatiotemporal trends in microclimates among the three
predominant mesohabitats of the Great Basin Scrub in the
Solar radiation,
wind speed and air temperature within each mesohabitat of this portion of the
Substratum temperature in the dappled/mixed
shade varies with time of day, microhabitat, and nanohabitat because of the
continuous change in the azimuth and declination of incoming solar radiation,
particularly where the sunlight strikes beneath the foliage-perimeter of the
shrubs. Differences in plant physiognomy
between S. vermiculatus and A. tridentata appear to affect the
consistency of the shading effect of the foliage appears to vary slightly. That
is, a deeper and more consistent shade may be produced by greasewood (S. vermiculatus). If so, it a reasonable speculation is that 1)
greater and spatially more even branching in the volume of foliage that
comprises the above ground vegetation and 2) shorter distance between the underside
of the foliage volume and substratum in S.
vermiculatus may produce the more consistent shading. The sun-shade patterns and the temperatures
in patches of sun and shade of various sizes under small and large shrubs of
both species yet need to be documented before the effects of the thermal
environments under these shrubs on lizards can be understood.
II. Copper model operative environmental
temperature
Averaged copper model
temperature measurements
The copper models absorbed heat via conduction
and radiation from the soil, foliage, and air. The time lag of the temperatures
recorded for copper lizard models and iButtons relative to the air and
substratum temperatures measured by the weather station are readily apparent in
Figure 4. The iButtons, however were recording temperature nearer
the substratum and midway between base and perimeter of the perennial, hence
the iButtons measured nanohabitat temperatures (e.g., nanohabitat and nanoclimate)
of dappled lighting, whereas the copper models measured a mix of sun-shade edge
and dappled lighting conditions nearer the plant perimeter. As a general trend, however, it appears that
the data from the copper lizard models data correlated with the iButton data.
These spatiotemporal similarities may be because both the iButtons and copper
models were integrating temperature from around them as well as the incoming
heat waves.
Variation in temperatures
among copper models with different body positions exposed to varying sunlight
levels and wind speeds.
Comparisons of
temperatures of the different lizard copper model body positions in open
microhabitats, exposed to full sunlight, and under varying levels of exposure
to showed consistent differences among models.
These results corroborate the findings of Muth’s study of
thermoregulation postures of Callisaurus
draconoides (Muth, 1977), a common prey species of G. wislizenii in the Mojave and
Copper model
applications in predicting G. wislizenii spatiotemporal activity
The data provided by the copper models in this
study can be directly applied towards predicting and explaining G.
wislizenii spatiotemporal behavior patterns. This confirms earlier, seminal
work with copper model (Bakken 1992).
And similar to a study of thermoregulation in another lizard, (Bauwens
et. al., 1996), a modestly strong inference from the preliminary results of
this study is that in the heat of the day (
III. Focal observations of Gambelia wislizenii behavior
Smaller lizards
with smaller body volume relative to body surface area than larger lizards can
warm more quickly in the morning sun, thereby perhaps becoming active earlier
in the day than larger lizards. This
same phenomenon, however, may cause an inverse relationship of body size and
rate of overheating when venturing into the sunlit open areas in late morning
and
Even
with the trough in their activity during
The
data from the copper models and the sun-shade patterns and substratum and above
ground temperatures associated with sun and shade under perennials must be
considered preliminary. Similarly, the
low sample sizes for each size-sex class for each period of the day render the
behavioral results suggestive, but not definitive. Moreover, a careful examination of arthropod
prey captured by Gambelia wislizenii,
relative to the availability of those prey has yet to be performed. The results of the work on lizard behavior in
relation to its thermal environment in the Alvord Basin, however, is a firm
foundation, providing the understanding needed for the next round of research
endeavoring to document, explain, and understand the spatiotemporal patterns of
desert reptiles.
Literature Cited
Abts, M. L. 1976. Thermal ecology and movement in the leopard lizard, Gambelia
wislizenii. Unpublished MA Thesis.
Anderson, R.A.
2006, in press. Food acquisition modes and habitat use in Lizards: questions
from an integrative perspective. In
Bakken G.S.
1992. Measurement and application of operative and standard
operative procedures in ecology. Am Zool 32:194-216.
Bauwens, D., P.E. Hertz, and A, Castilla. 1996. Thermoregulation in lacertid lizard:
the relative contributions of distinct behavioral mechanisms. Ecology 77: pp
1818-1830.
Grant, B. and A.E. Dunham. 1988. Thermally imposed time constraints on
the activity of the desert lizard sceloporus
merriami. Ecology: 69: pp 167-176.
Larson,
P. and L. Larson. 1977.
A Sierra Club Naturalist’s Guide to the Deserts of the
Southwest. Sierra Club,
Lappin,
A.K. and E. J. Swinney.
1999. Sexual dimorphism as relates to natural history of leopard lizards
(Crotaphytidae: Gambelia). Copeia 1999:649-660
Muth, A.
1977. Body
temperatures and associated postures of the zebra-tailed lizard, Callisaurus draconoides. Copeia
1: 122-125.
Muth, A.
1977. Thermoregulatory
postures and orientation to the sun: a mechanistic evaluation for the
zebra-tailed lizard, Callisaurus
draconoides. Copeia 4: 711-715.
Parker, W.
S. and E. R. Pianka.
1976. Ecological observations on the leopard lizard (Crotaphytus wislizenii)in
different parts of its range. Herpetologica 32: 95-114.
Rose, E.L. 2004. Foraging
behavior in Gambelia wislizenii the long-nosed leopard lizard, in
Sigler,
J.W. and W.F. Sigler.
1994. Fishes of the
Steffen, J. E. 2002. The ecological correlates of habitat use for he long nose leopard
lizard, Gambelia wislizenii, in
southeast
Tanner, W.
W. and J. E. Krogh.
1974. Ecology of the leopard lizard, Crotaphytus wislizenii at the
Tanner W. W
and J E. Krogh. 1974.Variations in activity as seen in four sympatric lizard
species of southern
Heath, J. E. 1965. Temperature regulation and diurnal activity in horned lizards.
Waldschmidt, S. 1980.
Orientation to the sun by the Iguana lizards Uta stansburiana and Sceloporus
undulatus: hourly and monthly variations. Copeia 3: 458-462.
Whitaker, J
and C. Maser. 1981. Food
habits of seven species of lizards from Malheur county, southeastern